10. El Fundamento de la Variación Heredable, Parte 2
3 de Mayo de 2013. Temas: Genética
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.
En este artículo tratamos sobre cómo la variación en el ADN se traduce en variación en la estructura y expresión de las proteínas.
En el artículo anterior discutíamos cómo la replicación del ADN es directamente facilitada por su estructura, ya que una mitad de la doble hélice de ADN puede servir como plantilla para hacer la otra mitad. También discutíamos cómo el ADN, aunque bien dotado para su papel hereditario, no lo está en absoluto para llevar a cabo funciones celulares, pero que las proteínas cubren esos papeles, esas funciones. Una vez cubiertos estos detalles podemos ya discutir cómo la información hereditaria del ADN es transformada en la diversidad funcional que observamos en las proteínas, y cómo la variación juega un papel en ese proceso. El primer paso en esta discusión nos obliga a investigar cómo funcionan los cromosomas y los genes.
Genética Molecular 102: Cromosomas y Genes
Los seres humanos tienen 46 cromosomas en cada una de sus células, y están dispuestos en pares. Recibimos uno de los cromosomas de cada par, de cada uno de nuestros padres: los óvulos contienen 22 cromosomas no sexuales más un cromosoma X, y los espermatozoides contienen 22 cromosomas no sexuales más un cromosoma que puede ser X o Y. Cada cromosoma es una larga doble hélice de ADN, con millones de pares de bases de ADN. Nuestros cromosomas más largos tienen unos 250 millones de pares de bases, y los más cortos unos 50 millones. En conjunto, el genoma humano tiene unos 3000 millones de pares de bases de ADN en cada juego de 23 cromosomas, es decir, un total de unos 6000 millones si contamos ambos juegos.
Los genes, las unidades de función biológica codificadas en nuestro ADN, se encuentran distribuidos en esos 23 pares de cromosomas. Como todos los buenos conceptos en Biología, lo que realmente constituye un “gen” es “confuso”, poco claro, pero para nuestros propósitos definiremos un gen como una secuencia de pares de bases de ADN de un cromosoma que se utiliza para formar un producto, funcional, que no es ADN. El hombre tiene unos 20 000 genes que pueden estar bastante repartidos por los cromosomas, con mucho más ADN, que no forma genes, entre ellos. Si representáramos el cromosoma mediante una línea negra continua, como suele hacerse en muchos textos de Genética, podríamos ampliarlos con un zoom para ver las características de alguno de sus muchos genes. En este caso, esto es un gen que forma una proteína:
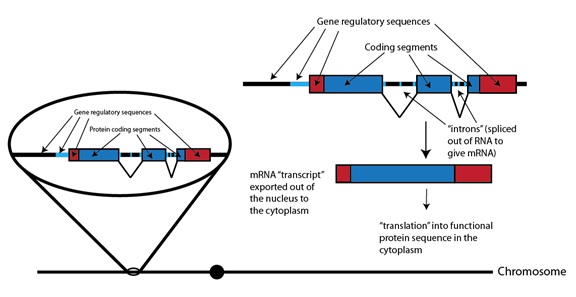
Secuencias reguladoras del gen
Segmentos que codifican proteína
“Traducción” en una secuencia de
proteína funcional, con un patrón alterado que produce la condrodisplasia
Secuencias reguladoras del gen
Segmentos codificantes
“intrones” (eliminados
en el ARN para
dar ARNm)
ARNm “transcrito”
exportado fuera del
núcleo al citoplasma
“Traducción” en una secuencia de
proteína funcional en el citoplasma
Cromosoma
(Nota: el “punto” en la línea que representa el cromosoma es el centrómero, una secuencia de ADN a la que algunos componentes celulares se unen para arrastrar a los cromosomas durante la división celular).
Para empezar, podemos ver que las partes del gen que son utilizadas para especificar la secuencia de aminoácidos de la proteína (las cajas azules) son sólo una parte del conjunto total. Otras secuencias, tales como las líneas de color azul claro y las cajas rojas, son secuencias que dirigen a ciertos tipos celulares para fabricar esa proteína, y para especificar cuánta tienen que fabricar. Todas las secuencias representadas como cajas son incluidas construyendo lo que se llama el “ARN mensajero”, o ARNm, que es como una versión de un tipo de ADN con una sola cadena, cuya longitud es sólo lo que mide la secuencia del gen y que, a menudo, incluye secuencias que intercalan las secciones que codifican la estructura de la proteína, los llamados “intrones”, que pueden verse en la figura anterior. Este ARNm “copia de trabajo” del gen se utiliza luego para dirigir la síntesis de la proteína a través de un proceso llamado traducción genética.
Si todo esto parece un poco complicado, no nos preocupemos; para nuestro objetivo aquí basta con reconocer que los genes son (a) una pequeña sección de una molécula de ADN, que es mucho más larga (es decir, el cromosoma); (b) tienen ciertas secuencias que determinan la secuencia de la proteína que codifican (es decir, el orden de sus aminoácidos), y (c) otras secuencias reguladoras que no son parte del propio código de la proteína, pero que funcionan más bien como señales para decirle a las células cuándo y dónde debe fabricarse, o “expresarse”, la proteína.
Introducir variación
Con estos detalles en mente, consideremos ahora cómo la variación a nivel del ADN puede afectar a la estructura del cromosoma. Como vimos antes, cuando se copian los cromosomas pueden cometerse errores de copia del ADN. No es de extrañar, pues, que muchos tipos de mutaciones puedan también afectar a la función de los genes y, en definitiva, a las características del organismo:
Mutaciones en un solo par de bases: el error en el emparejamiento de los ácidos nucleicos puede producir copias de cromosomas que difieren del original en un par de bases, como veíamos. Estas llamadas “mutaciones puntuales” pueden darse bien dentro de los genes, tanto en el ADN regulador como en el ADN que codifica proteínas, o bien en las secuencias que hay entre los genes. Los cambios de un solo par de bases en el ADN que codifica proteínas pueden no tener efecto ninguno en las proteínas, porque suele haber a menudo diferentes secuencias de ADN que producen la misma secuencia de aminoácidos, un carácter conocido como “redundancia” (o degeneración) del código genético. Otros cambios pueden alterar la secuencia de aminoácidos sustituyendo un aminoácido por otro, pero también pueden no tener efectos sobre la función de la proteína porque muchas funciones proteínicas pueden ser realizadas por secuencias proteínicas ligeramente diferentes. Otros cambios pueden reducir, o incluso anular la función de la proteína. Y finalmente, otros pueden mejorar la función de la proteína, dotarla de una mejor actividad enzimática, por ejemplo.
En el ADN regulador también son posibles los cambios, y los efectos de estos cambios también pueden ser neutros, perjudiciales o beneficiosos. Lo interesante del ADN regulador es que pequeños cambios pueden tener efectos bastante grandes sobre dónde y cuándo se forman las proteínas, y los cambios que afectan a genes clave que funcionan en el desarrollo temprano pueden tener efectos significativos que, en fases posteriores, afectan al organismo en su conjunto. Examinaremos esto con cierto detalle en posteriores artículos de esta serie.
Deleciones: a veces se pueden perder trozos de AND durante la replicación cromosómica, debido a procesos de rotura y de unión de nuevo. Las deleciones, a veces afectan sólo a unos pocos pares de bases, pero en ocasiones pueden abarcar miles de pares de bases. Se pueden perder partes de genes, o incluso genes enteros, y los genes que flanquean una deleción pueden aproximarse entre sí. Como hemos visto en las mutaciones puntuales, las deleciones pueden no tener efecto, pueden tenerlo perjudicial, o incluso ser beneficioso, dependiendo de cada caso. Por ejemplo, algunas veces las deleciones eliminan secuencias reguladoras que apagan la expresión génica en ciertas células. Al eliminar esa secuencia se permite al gen expresarse donde antes no lo hacía lo que, de nuevo, puede ser neutro, perjudicial o ventajoso, dependiendo de las circunstancias
Duplicaciones: son lo contrario que una deleción; una porción de la secuencia de un cromosoma se duplica y acaban quedando una al lado de la otra. Como en las deleciones, las duplicaciones pueden ser pequeñas o tener a una longitud de miles de pares de bases, abarcando muchos genes, y de igual modo pueden ser neutras, perjudiciales o ventajosas.
Un mecanismo común, que produce duplicaciones y deleciones simultáneamente, tiene lugar durante la recombinación en las células que van a formar los óvulos o los espermatozoides. Recordaremos que es “sobrecruzamiento” el término utilizado para describir la rotura física y la vuelta a unirse en los cromosomas, para “mezclar y emparejar” las secuencias entre los pares de cromosomas durante las divisiones celulares que conducen a la formación de los gametos (es decir la meiosis). Normalmente para este intercambio los cromosomas se emparejan, alineando sus secuencias, que son casi idénticas, y sufren una rotura y un empalme precisos inmediatamente:
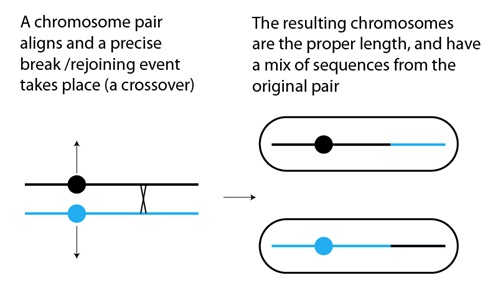
Una pareja de cromosomas
se alinea y luego tienen lugar
una rotura y un empalme posterior precisos (un sobrecruzamiento)
Los cromosomas resultantes
son de la longitud adecuada,
y tienen una mezcla de las secuencias
de la pareja original
Lo que puede ocurrir, con una frecuencia muy baja, es que los dos cromosomas no alineen sus secuencias correctamente. El alineamiento depende de que las mismas secuencias de uno y otro cromosoma se encuentren y se unan entre sí. Puede haber errores debidos a la repetición de secuencias entre genes, secuencias que “confunden” a los cromosomas, haciéndoles creer que han encontrado su secuencia correcta de alineamiento cuando, de hecho, hay dos lazadas de secuencias desemparejadas, una en cada cromosoma. Si tiene lugar un sobrecruzamiento entre esas dos lazadas, el resultado es un cromosoma con una duplicación, y otro con una deleción:
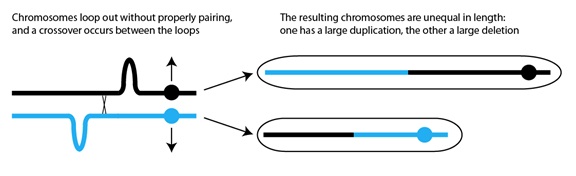
Los cromosomas forman lazadas
sin el emparejamiento adecuado,
y sucede el sobrecruzamiento
entre las dos lazadas
Los cromosomas resultantes
son de una longitud distinta:
uno tiene una gran duplicación,
y el otro una gran deleción
Desde luego, esta lista de tipos de mutación no es exhaustiva; por ejemplo, ya hemos visto cómo los elementos autónomos de ADN parásito llamados “transposones” pueden insertarse en los cromosomas, afectando a sus funciones o contribuyendo a funciones nuevas.
En resumen: constancia y cambio
En conjunto, estos mecanismos introducen la variación en las poblaciones y, como esta variación se da en el ADN, la variación es heredable. La variación a nivel cromosómico puede influir en la función de los genes y, en último término, en los rasgos del organismo. Los cambios a nivel del ADN que producen una variación con sentido a nivel del organismo quedan a disposición de la selección natural para que actúe sobre ellos; y ya hemos visto algunos ejemplos de mutaciones concretas, tales como la duplicación de los genes de la amilasa en el hombre y en el perro. Otras mutaciones, desde luego, son seleccionadas en contra y, con el tiempo, son eliminadas de las poblaciones. Las propiedades del ADN, tanto como agente de constancia y como cambio hereditario, significan que las poblaciones no son del todo genéticamente estables: pueden cambiar a lo largo del tiempo, aunque las características del ADN que le convierten en un medio muy preciso de transmisión de información aseguran que dichos cambios serán probablemente imperceptibles a nivel del organismo.
Como veremos en el siguiente artículo de esta serie, esta inestabilidad genética puede separar a distintas poblaciones de la misma especie en trayectorias diferentes, y permitir que se acumulen las diferencias que finalmente conducen a la formación de una nueva especie.


