06. Nuevos Genes, una Dieta Nueva, y sus Implicaciones en los Orígenes del Perro
5 de Abril de 2013. Temas: Genética, Historia de la Vida
Nota: Esta serie de artículos ha sido concebida como una introducción básica a la ciencia de la evolución para no especialistas. Aquí se puede ver la introducción a esta serie o volver al índice aquí.

En este artículo examinamos un caso de selección natural actuando durante el proceso de domesticación temprana, y cómo la selección artificial moldeó más tarde el genoma del perro durante la creación de las razas.
¿Es la selección artificial una analogía útil de la selección natural?
Recordaremos que Darwin presentaba los ejemplos de selección artificial como evidencia de que la selección natural podía producir el mismo efecto sobre las especies: un cambio en las características medias de una población a lo largo del tiempo. Darwin, por supuesto, no tenía ni idea de cómo funcionaba la herencia. Ahora que tenemos acceso a las secuencias del genoma, sin embargo, tenemos la oportunidad de comprobar la solidez de la analogía de Darwin a nivel molecular. Para ello, sin embargo, tenemos que comparar los detalles moleculares de ambos tipos de sucesos, de ambos tipos de selección. Y es interesante, que el genoma del perro también muestra señales de la selección natural durante el proceso inicial de domesticación, el cual podemos comparar con ejemplos de selección artificial.
¿Carne o patatas? La selección natural durante la domesticación temprana del perro
Precisamente el mismo estudio (que tratábamos en el artículo anterior) que demostraba que había una selección significativa de la variación en los genes relacionados con el sistema nervioso en el perro durante el proceso de domesticación temprana, también identificaba la selección sobre un tipo de genes implicados en el metabolismo del almidón. El almidón es una larga cadena de moléculas de azúcar (glucosa) unidas que las plantas utilizan como mecanismo de almacenamiento de energía. El lobo no tiene una dieta con grandes cantidades de almidón, pero sí ingiere algo en los frutos silvestres que a veces consume (por ejemplo, las bayas). Para sacar provecho del almidón, los mamíferos utilizan un tipo de enzimas, llamados amilasas, que rompen la cadena de almidón en moléculas separadas de glucosa. En el lobo, la amilasa se produce en el páncreas. Y como el lobo, el perro también tiene una enzima amilasa pancreática, pero en lugar de tener un solo gen, todos los perros modernos tienen entre 2 y 15 copias de ese gen, mientras que los lobos tienen sólo una. Las copias del perro se sitúan juntas en su genoma, justo al lado de donde el gen original de la amilasa se encuentra en el lobo, indicando que las copias del gen son duplicadas durante la replicación cromosómica. Estas copias extras del gen de la amilasa aumentan mucho la cantidad de enzima amilasa que los perros sintetizan, en relación con los lobos, y permiten que los perros se beneficien de una dieta mucho más rica en almidón de lo que los lobos podrían hacer.
Esta dieta es, desde luego, una dieta típicamente humana: el uso generalizado de plantas ricas en almidón es característico de la agricultura humana, que aparece en escena hace unos 10.000 años, más o menos. La asociación del perro con el hombre en ese momento parece, por tanto, que proporcionó una ventaja selectiva a los perros, que pudieron obtener más beneficios de la comida que recibían (o aprovechaban) de procedencia humana. En otras palabras, el ambiente en que se encontraban los perros (con acceso a alimentos con elevadas cantidades de almidón asociados a los asentamientos humanos) produjo la selección: los perros que pudieron obtener un aumento de las ventajas nutricionales del almidón serían capaces de reproducirse a un ritmo más elevado que los que no pudieron. Con el tiempo, la capacidad media de los perros para metabolizar el almidón haría crecer la población, a medida que cada vez más perros tuvieran duplicaciones. Como no hubo elección humana consciente para identificar a los perros con genes duplicados de amilasa, y seleccionarlos para la cría, porque dicha característica no resulta directamente observable, resulta que éste es un ejemplo de selección ambiental, de selección natural. Ni siquiera hoy día son capaces los humanos de seleccionar perros para mejorar el metabolismo del almidón, a pesar del hecho de que los perros modernos siguen mostrando variabilidad en cuanto al número de copias del gen de la amilasa que poseen.
Del perro al perro salchicha: la selección artificial de un nuevo gen
Tras domesticar al lobo en perro, no pararon nuestros esfuerzos selectivos, desde luego: los humanos utilizaron la selección artificial para crear más de 400 razas de perros, y podemos ver el resultado de esta selección sobre el genoma del perro al comparar unas razas con otras. Un ejemplo llamativo de diferencia entre las razas es la longitud de la pata. Algunas razas se definen por unas patas especialmente cortas, sin una reducción proporcional de su tamaño corporal, como los perros basset y los perros salchicha. Este rasgo, conocido técnicamente como “condrodisplasia”, es uno de los que se seleccionaron en base a su utilidad en determinados aspectos relacionados con la caza, tales como la persecución de animales excavadores. La base genética de este rasgo resulta ser otra duplicación que tuvo lugar hace tiempo en los perros domésticos: todas las razas de patas cortas comparten la misma innovación genética.
Nota: los detalles moleculares de esta duplicación son algo más complejos que la simple duplicación lateral del gen de la amilasa que acabamos de discutir. He incluido los detalles más abajo para aquellos lectores que quieran llegar a precisar más. La moraleja, sin embargo, es muy simple: en este caso, un rasgo nuevo (el acortamiento de las patas) se logró al duplicarse un gen y adquirir la copia del gen nuevas propiedades durante la propia duplicación. El rasgo de patas cortas se observó y fue seleccionado por los criadores humanos, convirtiéndolo en un ejemplo de selección artificial que condujo a nuevas razas con una característica específica. Si los detalles de la propia duplicación no son de nuestro interés, podemos pasarlos por alto y proseguir con la historia en el apartado “Comparar la selección artificial con la natural en el perro”.
La base molecular de cómo este nuevo gen responsable de la condrodisplasia llegó a aparecer es muy parecida a lo que ya hemos visto en detalle anteriormente: el nuevo gen es, en efecto, un pseudogen procesado que adquirió una función en el momento de su duplicación. Como la nueva copia fue funcional desde su aparición, no debe describirse como un pseudogen, sino más bien como un retrogen: esto es, la copia de ARNm de un gen que (a) ha sufrido una transcripción inversa de ARNm de nuevo a ADN, que (b) se ha insertado en el genoma a continuación de las secuencias que pudieran dirigir su expresión, y que (c) llegaron por selección a mantenerlo:
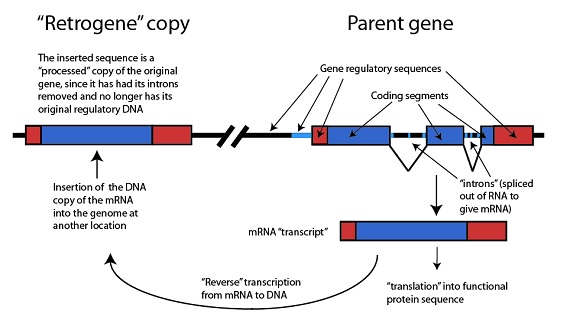
“Retrogen” copiado
La secuencia insertada es una copia “procesada” del gen original, porque se han quitado sus intrones y ya no tiene su ADN regulador original
Inserción de la copia de ADN del ARNm dentro del genoma en una nueva localización
Gen original parental
Secuencias reguladoras del gen
Segmentos codificantes
“intrones” eliminados en el ARN para dar ARNm (acido ribonucleico mensajero)
ARNm transcrito
“Traducción” en secuencia de proteína funcional
Transcripción “inversa” del ARNm en ADN
(Nota: DNA= ADN= Acido desoxirribonucleico; RNA= ARN= Acido ribonucleico
El nuevo retrogen resulta ser una copia del gen fgf4, un importante regulador del crecimiento y del desarrollo. Por razones que aún no han quedado claras, el retrogen fgf4 interfiere con el crecimiento normal del hueso y provoca el rasgo de patas cortas en los perros con condrodisplasia:
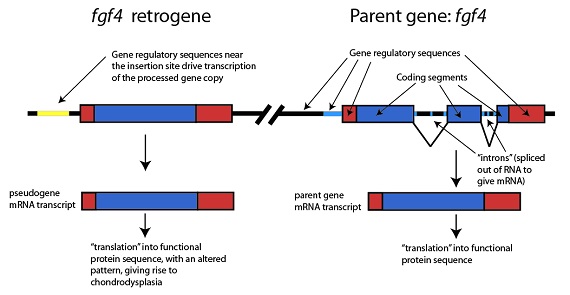
Retrogen fgf4
Secuencias reguladoras del gen
cerca del punto de inserción regulan
la transcripción de la copia “procesada”
del gen
ARNm transcrito
del seudogen
“Traducción” en una secuencia de
proteína funcional, con un patrón alterado que produce la condrodisplasia
Gen original: fgf4
Secuencias reguladoras del gen
Segmentos codificantes
“intrones” eliminados
en el ARN para dar ARNm
ARNm transcrito del gen original
“Traducción” en una secuencia deproteína funcional
Otra característica interesante del nuevo retrogen fgf4 es que su secuencia reguladora (mostrada en amarillo en esta figura) deriva de un transposón. Como hemos visto antes, los transposones son ADN autorreplicante parásito, autónomo, que pueden en ocasiones llegar a formar parte de los genes de sus hospedadores (y éste es otro ejemplo). El nuevo retrogen fgf4 se ha “pergeñado”, ensamblado, a partir de secuencias de transposones unidas a la secuencia codificante original fgf4, y adquiere una nueva función, la de reducir el crecimiento de la pata, que no aparece ni en la forma original del gen fgf4 ni en la secuencia de transposones.
Comparar la selección artificial con la natural en el perro
Mientras que la mutación que produjo el acortamiento de las patas en algunas razas de perros resultaba un ejemplo especialmente dramático de cómo surge una nueva variación (al implicar lo que supone, en efecto, el nacimiento de un nuevo gen), había otras muchas regiones genómicas seleccionadas durante la creación de las nuevas razas de perros. Hay muchos otros ejemplos más corrientes: el pequeño tamaño corporal común en el grupo de los “perros miniatura” se produce por la selección en la variación en el gen del factor de crecimiento insulínico tipo 1 (IGF1); se han identificado variaciones en tres genes claves como responsables de la variación en la coloración de la piel, e incluso se ha descrito la variación en un gen que produce la típica rugosidad de la piel de los perros Shar-Pei. En estos casos, no es la producción y selección de genes duplicados o de genes nuevos la que es responsable de esos rasgos, sino que más bien son las pequeñas mutaciones en los genes ya existentes las que alteran la función de dichos genes en comparación con su estado ancestral en los lobos o en los perros primitivos. Una vez más, la cuestión principal queda clara: pequeños cambios en el ADN, en combinación con la selección artificial, pueden llegar a suponer cambios morfológicos mucho mayores dentro de una población en un tiempo relativamente corto.
Así que, ¿hasta qué punto son comparables la selección natural y la artificial? Ambas formas de selección tienen la posibilidad de modificar, con el tiempo, las características medias de una población. Además, los fundamentos moleculares de las mutaciones en ambos tipos de selección son comparables: el rasgo debe surgir por una mutación que produzca una nueva variante heredable en la población. Pero ¡cuidado!: en el caso de la selección artificial, la inteligencia y la voluntad humanas no pueden producir la variación requerida, sino sólo seleccionarla positivamente una vez que haya aparecido. En ambos casos tenemos ejemplos de mutaciones en pequeñas secuencias, de duplicaciones e incluso de la aparición de genes nuevos. De hecho, la única diferencia radica en el paso de la selección: el filtro que posibilita que ciertas variantes se reproduzcan más que otras.
La acción humana podría ser una forma eficiente de selección, como lo es también la naturaleza. Por ello, el uso que hace Darwin de la selección artificial como evidencia de la selección en la naturaleza sigue siendo un enfoque válido, incluso a nivel molecular.
En el siguiente artículo de esta serie exploraremos cómo la selección natural ha moldeado la función de la amilasa tanto en el linaje humano como en los perros.
Lecturas complementarias:
Akey, J.M., et al.(2010). Tracking the footprints of artificial selection in the dog genome. PNAS 107; 1160 – 1165. (enlace)
Axelsson, E., et al. (2013). The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature 495; 360 – 364. (enlace)
Parker, H.G., et al. (2009). An Expressed Fgf4 Retrogene Is Associated with Breed-Defining Chondrodysplasia in Domestic Dogs. Science 325: 995 – 998. (enlace)


